25/04/2024 (2024-04-25)
[Source : viroliegy.com]
[Traduction : Jean Bitterlin]
L’Article d’Enders sur la rougeole (1954)
Par Mike Stone
L’article de John Franklin Enders de 1954 est considéré comme la preuve définitive de la découverte d’un « virus » de la rougeole. Cette preuve a été présentée comme l’« isolement » de la rougeole et a servi de base à l’élaboration du vaccin. Il a également jeté les bases de l’utilisation d’expériences de culture cellulaire comme moyen de cultiver des « virus », ainsi que des affirmations qui en découlent, selon lesquelles les preuves indirectes connues sous le nom d’effets cytopathogènes [ou cytopathiques] (ECP) peuvent servir de mesure de substitution pour les « virus » présents dans la soupe toxique de la culture cellulaire. La lecture de l’article et des conclusions d’Enders révèle cependant une histoire complètement différente de celle de l’isolement d’un « virus ». Voici les points saillants de son article :
Propagation dans les cultures de tissus d’agents cytopathogènes provenant de patients atteints de rougeole
« De nombreuses tentatives ont été faites dans le passé pour propager l’agent de la rougeole chez des animaux inférieurs, dans des embryons de poussins et dans des cultures de tissus (1-3). Les résultats des différents chercheurs étaient souvent divergents ou directement contradictoires. Il est cependant apparu assez clairement que les singes, en particulier M. Mulatta, sont modérément sensibles à l’inoculation expérimentale (3). En outre, les recherches de Rake, Shaffer et leurs collaborateurs ont fourni des preuves suggérant que l’agent qui passait à travers des filtres retenant les bactéries pouvait être maintenu indéfiniment lors de passages en série dans l’embryon de poussin en développement (4,s). Ces travailleurs (5) ont également confirmé les observations antérieures de Plotz (6) qui avait apparemment réussi à cultiver l’agent dans une culture cellulaire modifiée en suspension de tissus embryonnaires de poussins. Le passage de l’œuf dans les mains de Shaffer et de ses collaborateurs (7,8) a souvent semblé modifier la pathogénicité de l’agent pour l’homme, comme l’indique le développement d’une maladie légère et très modifiée à la suite de l’inoculation de matériel adapté à l’œuf à des enfants sensibles.Dans certains cas, cette maladie modifiée semble être suivie d’une résistance à la rougeole, comme l’indiquent les résultats d’une exposition ultérieure, naturelle ou artificielle, à la forme virulente de l’agent (9). Depuis 1943, date de la dernière communication de Rake et de ses collaborateurs, aucun progrès important n’a été réalisé dans l’étude de l’étiologie de la rougeole. Ce fait peut être attribué en grande partie à l’absence d’une méthode de laboratoire pratique pour démontrer la présence de l’agent qui n’induit aucun changement reconnaissable dans les œufs ou les cultures de tissus de poussins. De plus, les tentatives répétées de Shaffer (10) pour mettre en évidence une réaction sérologique, telle que la fixation du complément, en utilisant du matériel provenant d’embryons de poussins infectés, ont échoué. En conséquence, les seules techniques disponibles ont consisté à inoculer le virus à l’homme ou au singe. La première méthode n’est évidemment pas pratique à réaliser de manière routinière et la seconde est fastidieuse, coûteuse et souvent peu concluante en raison de la variation de la sensibilité individuelle.
Avec ces considérations à l’esprit, nous avons récemment tenté de cultiver l’agent de la rougeole dans des cultures de cellules humaines et de singes en utilisant des procédures appliquées avec succès à la propagation des virus de la poliomyélite (11-13). Dans le sang et les lavages de gorge de cas typiques de rougeole, ont été mis en évidence des agents qui peuvent être maintenus en passage sériel dans des cultures de tissus et qui induisent des changements cytopathiques distincts dans les cellules épithéliales rénales. Un certain nombre de preuves ont été accumulées indiquant que des anticorps spécifiques de ces agents se développent au cours de la maladie. Notre objectif est de décrire ici ces observations de manière préliminaire. Des preuves supplémentaires de la relation entre ces agents et la rougeole seront recherchées dans des enquêtes futures.
Matériels et méthodes. Collecte des échantillons
Des lavages de gorge, du sang veineux et des selles ont été prélevés chez 7 patients le plus tôt possible après l’établissement du diagnostic clinique de rougeole. Dans cinq cas, le moment où les échantillons ont été prélevés par rapport à l’apparition de l’exanthème est indiqué dans les histoires de cas décrites ci-dessous ou dans le tableau I. Lorsque les patients en étaient capables, on leur a demandé de se gargariser avec 10-15 ml de lait écrémé neutralisé stérile. Certains échantillons de la gorge des jeunes enfants ont été prélevés à l’aide d’un coton-tige préalablement humidifié dans du lait. Après avoir frotté la gorge, l’écouvillon a été immergé dans 2 ml de lait. De la pénicilline, 100 u/ml, et de la streptomycine, 50 mg/ml, ont été ajoutées à tous les échantillons prélevés dans la gorge qui ont ensuite été centrifugés à 5450 tours/minute pendant environ une heure. Le liquide surnageant et le sédiment remis en suspension dans un petit volume de lait ont été utilisés comme inocula séparés dans différentes expériences, dans des quantités variant de 0,5 ml à 3,0 ml. Immédiatement après le prélèvement environ 10 ml de sang ont été placés dans des tubes contenant 2 ml d’une solution d’héparine à 0,05 %. Des quantités variant de 0,5 ml à 2,0 ml de sang total ont été utilisées comme inocula pour les cultures de tissus. Après addition d’antibiotiques comme décrit ci-dessus, des suspensions fécales à 10 % ont été préparées en broyant le matériel dans un milieu de liquide amniotique bovin. Les suspensions ont ensuite été centrifugées à 5450 tours/minute pendant environ une heure et les liquides surnageants ont été utilisés comme inocula, dans des quantités variant de 0,1 ml à 3 ml. Tous les échantillons ont été réfrigérés dans de l’eau et de la glace ou maintenus au froid à environ 5 °C depuis le moment de la collecte jusqu’à leur ajout aux cultures. Le temps maximum qui s’est écoulé entre la collecte des échantillons et l’inoculation a été de 35 heures ».
Techniques de culture des tissus
« Lors des premières tentatives d’isolement, on a utilisé des cultures en tubes à rouleaux1 (1 1 12) de rein humain, de poumon embryonnaire humain, d’intestin embryonnaire humain, d’utérus humain et de testicule de singe rhésus. Des passages ultérieurs des agents isolés ont ensuite été tentés dans le rein humain, la peau et le muscle embryonnaires humains, le prépuce humain, l’utérus humain, le rein de singe rhésus et le tissu embryonnaire de poussin. Des cultures stationnaires préparées selon la technique de Youngner (13) avec des reins humains et de singe rhésus tryptisinés ont ensuite été utilisées pour l’isolement des agents et leur passage. Le milieu de culture est composé de liquide amniotique bovin (go%), d’extrait d’embryon de bœuf (50/0), de sérum de cheval (5 %), d’antibiotiques et de rouge de phénol comme indicateur du métabolisme cellulaire (1 2). L’inhibiteur de trypsine de soja a été ajouté à ce milieu, sauf s’il a été utilisé pour la culture de reins humains et de reins de singe (11). Les fluides étaient généralement changés à des intervalles de 4 à 5 jours. Pour l’examen histologique, la croissance cellulaire, après avoir été fixée dans du formol à 10 %, a été incluse dans du collodion, déshydratée et colorée à l’hématoxyline et à l’éosine ».
Mode de passage en culture de tissus
« Le passage en série des différentes souches (tableau I) a été effectué comme d’habitude en retirant le milieu de culture entre le 4e et le 16e jour après l’inoculation et en transférant immédiatement 0,1 ml dans chacune d’un certain nombre de cultures fraîches. Le passage réussi de l’agent avec des fluides préalablement centrifugés à 2500 tours/minute pour éliminer les éléments cellulaires a également été démontré à plusieurs reprises. Des inocula plus importants (jusqu’à 1,0 ml) ont souvent été utilisés au cours des premières expériences avant que la résistance de l’agent au stockage à différentes températures n’ait été déterminée ».
Expérience. Changements cytopathiques induits par des agents isolés de cas de rougeole
« Le premier des 8 agents obtenus à partir du sang ou des lavages de gorge des cas de rougeole et présentant des propriétés comparables a été isolé dans des cultures de tissus rénaux humains après addition du sang du cas 3. Dans chacune des 3 cultures inoculées, des changements cytopathiques ont été observés au 7e jour. Comme ces changements présentaient une apparence caractéristique qui n’avait pas été associée jusqu’à présent à un virus, ils ont fourni les moyens de poursuivre l’étude de cet agent ainsi que d’autres qui ont été récemment isolés ».
« L’examen des matériaux colorés a également révélé des changements significatifs dans les noyaux des cellules géantes qui n’étaient pas visibles dans les préparations fraîches. Il s’agit d’une redistribution de la chromatine qui finit par prendre une position marginale où elle forme un anneau ou un croissant dense qui marche intensément avec le colorant basique. Parallèlement, la partie centrale du noyau est occupée par une substance apparemment homogène, de caractère acidophile, qui se rapproche étroitement de l’anneau de chromatine. Étant donné que dans ces préparations et dans d’autres qui ont été examinées par la suite, aucune zone claire non colorée n’a été observée entre la chromatine et cette masse acidophile, on ne peut pas affirmer que cette dernière représente un corps d’inclusion intranucléaire du type de celui qui est associé de manière caractéristique aux infections virales. Néanmoins, pour autant qu’on puisse le déterminer maintenant, sa présence ainsi que la margination2 de la chromatine constituent un critère utile d’infection pour les agents étudiés. Il faut cependant souligner que les changements décrits ci-dessus se produisent dans des cultures qui ont été incubées pendant des périodes relativement longues (par exemple 14-21 jours). Lorsque l’intervalle entre l’inoculation de l’agent et l’examen des cellules colorées (par exemple 4 jours) est plus court, la margination de la chromatine peut être incomplète ou inapparente et la substance acidophile peut n’être visible que sous forme de petites masses arrondies réparties çà et là parmi des matériaux nucléaires qui se rapprochent de l’arrangement normal ».
Domaine cytopathogène
« Le rein de singe est le seul autre tissu utilisé qui ait donné lieu à une croissance cellulaire dans laquelle les changements caractéristiques décrits ci-dessus ont été observés avec certitude après inoculation du virus. Dans des cultures composées en grande partie de cellules épithéliales rénales de singe, préparées selon la technique de Dulbecco modifiée par Youngner (13), on a régulièrement observé des changements cytopathiques qui ressemblent beaucoup à ceux produits par ces agents dans les cellules rénales humaines, comme on a pu le voir dans des préparations fraîches et colorées. Ces effets ont suivi l’addition de sang ou de lavages de gorge provenant de cas de rougeole, ainsi que de liquides de culture de tissus infectés provenant de passages antérieurs. Les cultures de reins de singe peuvent donc être appliquées à l’étude de ces agents de la même manière que les cultures de reins humains. Ce faisant, il faut toutefois garder à l’esprit que les effets cytopathiques qui ressemblent superficiellement à ceux résultant de l’infection par les agents de la rougeole peuvent éventuellement être induits par d’autres agents viraux présents dans le tissu rénal de singe (cf. dernier paragraphe sous G) ou par des facteurs inconnus. Dans quelques cultures de tissu de prépuce humain inoculé avec l’un des agents de la rougeole, des changements ressemblant à ceux observés dans les cellules rénales ont été notés dans l’excroissance épithéliale autour de certains fragments. Des observations supplémentaires seront toutefois nécessaires avant de pouvoir affirmer avec certitude que les cellules épithéliales dermiques sont spécifiquement attaquées par ces virus. Dans une expérience unique, aucune manifestation cytopathique n’a été observée pendant une période de 31 jours après l’inoculation de liquide de culture tissulaire infecté dans des cultures de peau et de muscle embryonnaires humains, de tissu utérin humain ou de tissu embryonnaire de poussin. Les tests visant à détecter la présence d’antigènes fixant le complément dans les liquides prélevés sur les cultures le 31e jour se sont révélés négatifs. Ces résultats sérologiques suggèrent qu’il n’y a pas eu de croissance du virus, puisque, comme on le verra par la suite, l’antigène semble se développer régulièrement après plusieurs jours dans les cultures de tissus rénaux infectés par le virus ».
Autres agents isolés au cours de cette étude
« Deux agents ont été isolés au cours de cette étude et ne semblent pas avoir de rapport avec ceux que nous venons de décrire. Le premier a été récupéré à partir des lavages de gorge d’un cas typique de rougeole survenu à l’école des garçons. Son large spectre cytopathogène, le caractère des changements cytopathiques induits et le fait que son infectiosité pour les cultures de tissus ait été neutralisée par le sérum de lapin immunisé contre l’herpès simplex ont permis de définir sa nature. Un deuxième agent a été obtenu à partir d’une culture non inoculée de cellules rénales de singe. Les changements cytopathiques qu’il induit dans les préparations non colorées ne peuvent être distingués avec certitude des virus isolés de la rougeole. Mais lorsque les cellules des cultures infectées ont été fixées et colorées, leur effet a pu être facilement distingué, car les changements internucléaires typiques des agents de la rougeole n’ont pas été observés. En outre, comme nous l’avons déjà indiqué, les liquides provenant de cultures infectées par l’agent n’ont pas réussi à fixer le complément en présence de sérum de rougeole convalescente. Il est évident que la possibilité de rencontrer de tels agents dans les études sur la rougeole doit être constamment gardée à l’esprit.
Discussion
Parmi les nombreuses expériences qui ont été rapportées dans le passé et qui décrivent l’isolement réussi de l’agent étiologique de la rougeole, seules celles dans lesquelles des singes ont été utilisés comme animaux de laboratoire ont été systématiquement confirmées par d’autres chercheurs. Il convient donc d’être très prudent dans l’interprétation de toute nouvelle affirmation selon laquelle le virus a été propagé chez d’autres hôtes ou dans d’autres systèmes. En conséquence, les résultats résumés ici doivent être soumis à l’analyse la plus critique.
Les faits suivants tendent à confirmer l’hypothèse selon laquelle les virus que nous avons décrits sont responsables de la maladie. Des agents transmissibles expérimentalement, présentant un effet cytopathogène similaire et caractéristique sur des cultures de cellules épithéliales humaines ou simiennes, ont été isolés à partir du sang ou des lavages de gorge de 5 des 7 cas typiques de rougeole au cours de la phase aiguë précoce. Un agent a été mis en évidence dans le sang de 4 des 5 cas dont les échantillons ont été prélevés et examinés par la méthode de la culture tissulaire. Ces résultats semblent particulièrement importants, car il est peu probable que des virus non liés à la rougeole soient régulièrement présents dans le sang circulant de ces personnes, dont certaines étaient géographiquement très éloignées les unes des autres.
Les changements pathologiques induits par les agents dans les cellules épithéliales en culture de tissus ressemblent, au moins superficiellement, à ceux observés dans certains tissus au cours de la phase aiguë de la rougeole. Si rien ne permet de conclure que les facteurs in vivo sont les mêmes que ceux qui sous-tendent la formation de cellules géantes et les perturbations nucléaires in vitro, l’apparition de ces phénomènes dans les cellules en culture est cohérente avec les propriétés qui pourraient a priori être associées au virus de la rougeole.
L’apparition, au cours de la maladie, d’anticorps capables de supprimer l’effet cytopathogène et de fixer le complément en présence de liquides de culture de tissus infectés constitue une preuve supplémentaire de l’étroite association des agents avec la rougeole. Il est évident qu’il est souhaitable d’obtenir des données supplémentaires à partir de tests effectués sur des sérums provenant d’un grand nombre de cas de rougeole ainsi que d’autres maladies infectieuses, en particulier les exanthèmes communs, afin d’éliminer tout doute subsistant concernant la spécificité de ces réactions sérologiques. L’accumulation de ces données est en cours.
Bien que nous ayons déjà obtenu des preuves indirectes considérables du rôle étiologique de ce groupe d’agents dans la rougeole, il reste à réaliser deux expériences essentielles pour établir cette relation. Il s’agit de produire la rougeole chez le singe et chez l’homme avec du matériel de culture tissulaire après un nombre de passages in vitro suffisant pour éliminer tout virus introduit dans l’inoculum d’origine. La récupération du virus de la maladie expérimentale chez ces hôtes devrait ensuite être réalisée.
Conclusion
Les résultats qui viennent d’être résumés soutiennent la présomption que ce groupe d’agents est composé de représentants de l’espèce virale responsable de la rougeole ».
https://doi.org/10.3181%2F00379727-86-21073
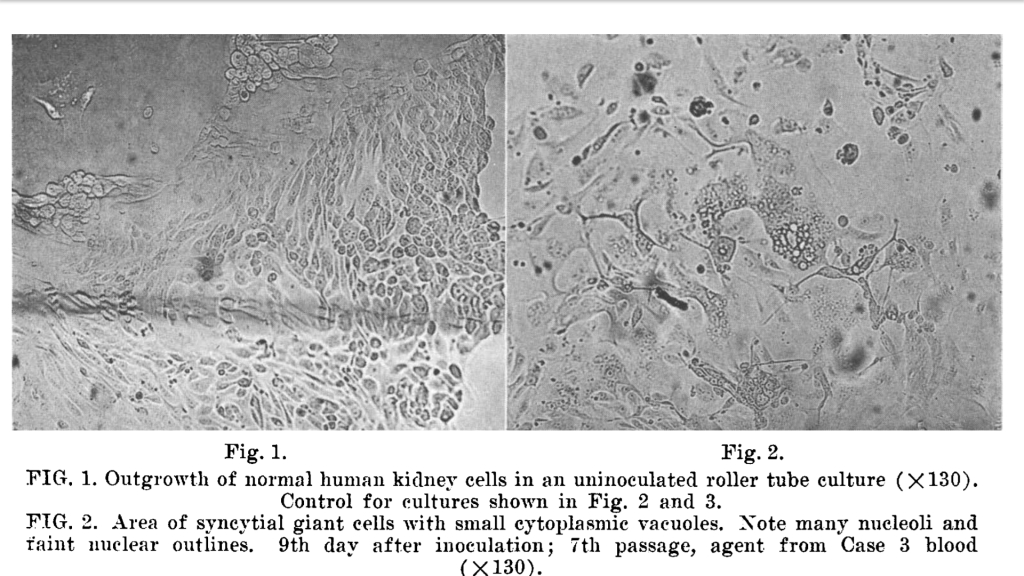
Légende des photos ci-dessus :
Figure 1 : Croissance de cellules rénales humaines normales dans un tube à rouleau inoculé (x 130). Contrôle pour les cultures présentées dans les figures 2 et 3.
Figure 2 : Zone de cellules géantes syncytiales avec de petites vacuoles cytoplasmiques. Notez la présence de nombreux nucléoles et de faibles contours nucléaires. 9e jour après inoculation ; 7e passage, agent du sang du cas 3 (x 130)
En résumé :
- Les résultats initiaux des différents chercheurs concernant la rougeole étaient souvent divergents ou carrément contradictoires.
- Enders a affirmé que les singes, en particulier les macaques rhésus, sont modérément sensibles à l’inoculation expérimentale.
- Shaffer et ses collaborateurs ont fourni des preuves suggérant que l’agent qui passait à travers des filtres retenant les bactéries pouvait être maintenu indéfiniment lors de passages en série dans l’embryon de poussin en développement.
- Cependant, le passage de l’œuf entre les mains de Shaffer et de ses collaborateurs semble régulièrement altérer la pathogénicité de l’agent pour l’homme, comme l’indique le développement d’une forme légère et très modifiée de la maladie à la suite de l’inoculation de matériel adapté à l’œuf à des enfants prédisposés.
- Les travaux susmentionnés de Shaffer semblent indiquer que le type de culture (tissu, cellule, embryon) a un impact profond sur les résultats obtenus des expériences.
- Depuis 1943, la recherche sur la rougeole n’a pas progressé, ce qu’Enders attribue à l’absence d’une méthode de laboratoire pratique pour démontrer la présence de l’agent qui n’induit aucun changement reconnaissable dans les œufs ou les cultures de tissus de poussins.
- Les tentatives répétées de Shaffer pour démontrer une réaction sérologique, telle que la fixation du complément, à l’aide de matériel provenant d’embryons de poussins infectés ont échoué.
- Enders est donc intervenu avec ses travaux pour affirmer qu’il avait été démontré que des agents [infectieux ou pathogènes] pouvaient être maintenus malgré le passage par une sériede cultures de tissus et qu’ils induisaient des changements cytopathiques distinctifs dans les cellules épithéliales rénales.
- Les patients ont été invités à se gargariser avec 10 à 15 ml de lait écrémé stérile.
- Certains échantillons de la gorge des jeunes enfants ont été prélevés à l’aide d’un coton-tige préalablement humidifié dans du lait.
- Après l’écouvillonnage de la gorge, l’écouvillon a été immergé dans 2 ml de lait.
- De la pénicilline 100 u/ml et de la streptomycine 50 mg/ml ont été ajoutées à tous les échantillons prélevés dans la gorge, qui ont ensuite été centrifugés à 5450 tours/minute pendant environ une heure.
- Le liquide surnageant et le sédiment remis en suspension dans un petit volume de lait ont été utilisés comme inocula séparés dans différentes expériences, dans des quantités variant de 0,5 ml à 3,0 ml.
- Environ 10 ml de sang immédiatement après le prélèvement ont été placés dans des tubes contenant 2 ml d’une solution d’héparine à 0,05 %.
- Après l’ajout d’antibiotiques comme décrit ci-dessus, des suspensions fécales à 10 % ont été préparées en broyant le matériel dans un milieu de liquide amniotique bovin.
- Des reins humains et de singe rhésus trypsinisés ont ensuite été utilisés pour « isoler » les agents et les faire passer.3
- Le milieu de culture était composé de :
- Liquide amniotique bovin ;
- Extrait d’embryon de bœuf ;
- Sérum de cheval ;
- Antibiotiques ;
- Rouge de phénol comme indicateur du métabolisme cellulaire.
- L’inhibiteur trypsine de soja a été ajouté à ce milieu, sauf lorsqu’il a été utilisé pour la culture de reins humains et de reins de singe.
- Le passage en série des différentes souches a été effectué en routine en retirant le milieu de culture entre le 4e et le 16e jour après l’inoculation et en transférant immédiatement 0,1 ml dans chacune des cultures fraîches.
Articles connexes sur les effets du passage sur la culture :
Sub-Culturing and Cell Culture Adaptations
Passage in Vero Cells: The Variant Game
- Enders a observé des effets cytopathogènes (ECP) après 7 jours de culture et a déterminé que ces changements présentaient un aspect caractéristique qui n’était pas associé de façon certaine à un « virus » ; il a donc supposé qu’il s’agissait d’une preuve de la présence de la rougeole dans la culture cellulaire.
- Au microscope électronique, étant donné que dans ces préparations et dans d’autres qui ont été examinées par la suite, aucune zone claire non colorée n’a été observée entre la chromatine et cette masse acidophile, on peut affirmer que cette dernière représentait un corps d’inclusion intranucléaire du type de ceux qui sont généralement associés aux infections « virales ».
- Cependant, Enders a déclaré qu’il peut être utilisé comme critère pour déterminer l’infection.
- Il a ensuite déclaré qu’il fallait souligner que les changements décrits ci-dessus se produisent dans des cultures qui ont été incubées pendant des périodes relativement longues (par exemple, 14 à 21 jours).
- Lorsque l’intervalle entre l’inoculation de l’agent et l’examen des cellules colorées (par exemple 4 jours) est plus court, la margination4 de la chromatine peut être incomplète ou inapparente.
- Une fois de plus, cela montre que les conditions de culture et la durée influencent grandement l’interprétation des résultats obtenus.
- Le rein de singe est le seul autre tissu utilisé qui a donné une croissance de cellules dans lesquelles les changements caractéristiques décrits ont été définitivement observés après l’inoculation du « virus ».
- Cependant, en ce qui concerne l’ECP observé, Enders a admis que les effets cytopathiques qui ressemblent superficiellement à ceux résultant de l’infection par les agents de la rougeole peuvent éventuellement être induits par d’autres agents « viraux » présents dans le tissu du rein de singe ou par des facteurs inconnus, montrant ainsi que non seulement l’ECP n’est pas spécifique, mais qu’il peut être causé par des facteurs autres qu’un « virus ».
Articles connexes sur d’autres facteurs pouvant causer l’EPC :
The Cytopathic Effect
Creating the Cytopathic Effect
- Deux autres agents ont également été « isolés » pendant que les travaux étaient en cours et, selon Enders, ils ne semblaient pas liés à ceux qu’il a décrits.
- Les changements cytopathiques induits dans les préparations non colorées ne pouvaient pas être distingués avec certitude des « virus » isolés de la rougeole.
- Enders a déclaré qu’il fallait évidemment garder constamment à l’esprit la possibilité de rencontrer de tels agents dans les études sur la rougeole.
- Cela démontre une fois de plus que le matériel avec lequel Enders a travaillé n’a pas été correctement purifié et que le « virus » de la rougeole n’a jamais été séparé du reste, étant donné la présence d’autres « agents ».
- Enders a déclaré qu’il fallait être très prudent dans l’interprétation de toute nouvelle affirmation selon laquelle le « virus » s’est propagé dans d’autres hôtes ou d’autres systèmes que les singes
- Il a également déclaré que les résultats qu’il a résumés doivent être soumis à l’analyse la plus critique.
- Enders estime que les preuves indirectes qu’il a présentées étayent l’hypothèse selon laquelle les « virus » qu’il a décrits sont responsables de la maladie.
- Enders a ensuite tiré à boulets rouges sur ses propres preuves cytopathogéniques en déclarant que si rien ne permet de conclure que les facteurs in vivo (dans un organisme vivant) sont les mêmes que ceux qui sous-tendent la formation de cellules géantes et les perturbations nucléaires in vitro (en dehors de l’organisme vivant), l’apparition de ces phénomènes dans des cellules cultivées est compatible avec les propriétés qui pourraient a priori être associées au « virus » de la rougeole.
- Même s’il s’est appuyé sur les anticorps pour démontrer qu’il avait « isolé » un « virus » de la rougeole, Enders a déclaré que d’autres tests étaient souhaitables pour éliminer tout doute subsistant concernant la spécificité de ces réactions sérologiques.
- Enders a admis avoir obtenu un nombre considérable de preuves indirectes.
- Il a également admis qu’il restait deux critères à réaliser et à remplir :
- La production de la rougeole chez le singe et chez l’homme avec du matériel de culture tissulaire après un nombre de passages in vitro suffisant pour éliminer tout « virus » introduit dans l’inoculum d’origine.
- La récupération du « virus » de la maladie expérimentale chez ces hôtes devrait alors être accomplie.
- Enders a conclu que ses preuves indirectes appuyaient la présomption que ce groupe d’agents est composé de représentants de l’espèce « virale » responsable de la rougeole.

[Voir : C’est l’heure du conte « Gain de Fiction »* avec RFK Jr. et ses amis !]
Il est clair, à la lecture de ce document, que l’on ne peut prétendre que le « virus » de la rougeole a été purifié/isolé, car les échantillons originaux ont subi de multiples altérations et ajouts. Pour commencer, les prélèvements de gorge ont été immédiatement placés dans du lait ou ont été effectués à l’aide de cotons-tiges imbibés de lait. Des antibiotiques ont ensuite été ajoutés à ces prélèvements de lait. Les échantillons de sang ont été placés dans des tubes contenant de l’héparine, qui est toxique pour les cellules. Des échantillons de matière fécale ont été broyés et ajoutés au sérum de liquide amniotique bovin. De nombreux produits chimiques/composés ont été utilisés au cours du processus d’« isolation ». Des cellules rénales humaines ou de singe trypsinisées ont été utilisées et il a été démontré que la trypsine a des effets négatifs sur les reins et qu’elle peut contenir de nombreux contaminants. Du liquide amniotique bovin, de l’extrait d’embryon de bœuf, du sérum de cheval, des antibiotiques, du rouge de phénol et, dans certains cas, des inhibiteurs de trypsine de soja ont été ajoutés à la culture. L’ajout de divers produits chimiques et composés altérant les cellules, d’ADN animal, etc. est l’exact opposé de la purification/isolation.
Les changements cytopathiques observés par Enders, qui l’ont amené à conclure à la présence d’un « virus », se sont produits après 7 jours de culture. Enders a déclaré que les changements cytopathiques (ECP) observés dans les cellules rénales de singe ressemblent à ceux des cellules rénales humaines et qu’elles peuvent être utilisées à la place des cellules humaines pour la culture, comme elles le sont régulièrement aujourd’hui. Cependant, le simple fait d’observer un ECP dans une culture cellulaire ne signifie pas qu’un « virus » est présent ni qu’il doit être tenu pour responsable de l’effet. N’importe lequel des composés toxiques ajoutés à la culture aurait pu à lui seul provoquer ces changements. Il n’y a aucune raison de supposer qu’un « virus » est à l’origine de la mort cellulaire. Enders admet même que l’ECP qu’il a observé n’est peut-être pas dû à un « virus » de la rougeole, mais qu’il pourrait être le résultat de différents « virus » ou d’autres facteurs encore inconnus. Ainsi, sa preuve essentielle n’était en fait pas une preuve du tout.
Enders a également admis que d’autres agents avaient été « isolés » en même temps que la rougeole et qu’ils ne pouvaient distinguer aucune différence entre l’ECP qu’ils prétendent être causé par un « virus » de la rougeole et l’ECP produit par l’un des autres agents. C’est une preuve supplémentaire qu’il ne s’agissait pas d’échantillons purifiés/isolés et qu’ils supposent que le « virus » qu’ils croient présent doit adhérer à certaines modifications de l’ECP, même s’il existe d’autres agents qui peuvent produire le même effet. L’ECP observé n’est pas spécifique à un « virus », quel qu’il soit5.
Enders a également fait quelques révélations surprenantes dans son document, qui tendent à faire tomber ses propres preuves. Tout d’abord, il prétend qu’il n’y a aucune raison d’affirmer que ce qui se passe IN VITRO (dans un laboratoire) a une quelconque relation avec ce qui se passe IN VIVO (au sein d’un organisme vivant). Or, tout ce qu’il a fait et les preuves qui en résultent, selon lui, proviennent d’expériences réalisées en dehors d’un organisme vivant.
Deuxièmement, Enders admet qu’il n’a recueilli que des preuves INDIRECTES du rôle des agents qu’il a étudiés, mais que d’autres expériences étaient nécessaires pour prouver qu’il avait effectivement obtenu6 un « virus » de la rougeole. Il fallait notamment vérifier si l’agent pouvait produire la rougeole chez l’homme ou le singe et si le « virus » de la rougeole pouvait être récupéré chez l’un ou l’autre. Ce document n’est donc pas du tout une preuve de l’existence d’un « virus » de la rougeole. Si cela ne suffisait pas à rendre ce point absolument clair, la conclusion d’Enders le fera certainement :
« Les résultats qui viennent d’être résumés soutiennent la présomption que ce groupe d’agents est composé de représentants des espèces virales responsables de la rougeole ».
Il s’agit de l’étude de référence sur la rougeole et elle n’apporte aucune preuve de l’existence d’un « virus » de la rougeole. Il présume (c’est-à-dire qu’il suppose vrai sans preuve) l’existence d’un « virus ». Tout ce qui a été construit sur la base de cet article frauduleux est donc également frauduleux, ce qui est la nature même de la « virologie » et de la « science » aujourd’hui.
Traduction Jean Bitterlin, 24 avril 2024
⚠ Les points de vue exprimés dans l’article ne sont pas nécessairement partagés par les (autres) auteurs et contributeurs du site Nouveau Monde.


















